
С.Т. Захидов
Московский государственный университет им. М.В.Ломоносова
В статье речь идет о явлении мутаций — аналогах термодинамических флуктуаций и их роли в процессах самоорганизации и развитии наследственной и биологической материй. Рассмотрены некоторые положения рапопортовской генетической теории в контексте с современными синергетическими концепциями.
Так кто ж ты, наконец?
Я — часть той силы, что вечно хочет
зла и вечно совершает благо
Гете «Фауст» (подстрочный перевод М.А.Булгакова)
…но и чрез сии разрушения
видна цель творения
Гердер
Любое отклонение от идеального случая
следует считать несравненно большей
ценностью, чем рядовой, в массовом
порядке ожидаемый, поскольку попутно
с возникновением отрицательной единицы
стоит пролагающая новый путь
положительная.
И.А.Рапопорт
Генетика и синергетика
Раз возникнув как самостоятельный раздел естествознания, генетика — экспериментальная наука о дискретной наследственности и изменчивости — динамично развивалась, не зная периодов упадка и застоя. В течение последних ста лет исследования в области генетики по значимости и интенсивности не уступали исследованиям в области квантовой физики. Овладение энергией гена — задача грандиозная, сопоставимая с решением проблемы управляемого термоядерного синтеза.
Современная генетика изучает материальную основу аппарата наследственности, манипулирует генетическими телами. Она также изучает мутационный процесс — процесс хаотизации генетического мира.
Синергетика — новая система познания мира в его единстве, глобальная наука, жадно изучающая все существующее во всей его сложности. Синергетика зародилась в недрах математики и теоретической физики, как область знаний о нелинейных процессах в различных средах. Сегодня она включает в себя идеи классической и неклассической (неравновесной) термодинамики, теорию хаоса и теорию катастроф, предметом ее изучения являются кооперативные явления, механизмы пространственно-временной самоорганизации и развития многообразных структур и подсистем в системах, бросающих вызов мировой энтропии.
Синергетика ведет поиск параллелей (образцов подобия) между явлениями живой и мертвой природы с целью установления универсальных закономерностей в эволюции материального мира. Она широко использует аппарат аналогий как инструмент для открытий и соединения различных ступеней Природы.
Синергетика, как когда-то атомистика, призвана создать творческую основу для нового революционного прорыва в естествознании и философии, сыграть роль нити Ариадны, позволяющей идти по правильному пути.
Синергетические теории дают нам новые представления об энтропии как реальном, могущественном факторе, действующем во всех уголках материального мира, напоминают, что энтропия не может быть «фигурой умолчания, а тем более огульного отрицания», а должна стать предметом глубокого анализа, что понятие энтропии применимо к широкому кругу явлений, процессов, систем, ко всем формам энергии, в том числе и абстрактным. Энтропия — это не только непроизводительные, необратимые потери энергии, отклонения от закономерных норм, но и фактор, способствующий упорядочению и совершенствованию сложных систем, их эволюции и развитию (Пригожин, Стенгерс, 2000а,б, Николис, Пригожин, 2003, Хакен, 1999, 2000, Климонтович, 2000а,б, Руденко, 2000, 2001, Рапопорт, 1965, 1972, 1993).
Несмотря на то что синергетика как самая активная методология, проникающая сегодня повсюду, и ничего при этом из науки не вытесняющая, сделала предметом своего изучения все природные системы, генетика не включена пока в круг ее интересов. Но как только между ними установится мощная связь, многое, что сегодня составляет содержание науки о наследственности и изменчивости, может испытать новое рождение.
В свою очередь, синергетика, если глубоко проникнет в сущность явлений и законы генетики, имеющей свои существенные теоретические отношения с квантовой механикой и химией, получит новые модели и плодотворные идеи, увеличит сумму знаний, увидит недостающие звенья. Она также получит верные ключи к пониманию тонких и, возможно, весьма специфических механизмов самоорганизации, и на этой почве — как шел эволюционный процесс, именно — от простого ли к сложному, или от сложного к сложному или, как предполагал Бэтсон, «…все существующее было когда-то предвечно создано во всей своей сложности и потом только изменялось, теряя по пути часть своих первозданных свойств или перетасовывая их в новых комбинациях».\
Исключительную роль в развитии и обогащении синерегетических теорий могут сыграть фундаментальные, теоретические исследования выдающегося советского ученого, основоположника химического мутагенеза, отца генетической термодинамики и генетической атомистики, автора современной теории генетической и биологической эволюции Иосифа Абрамовича Рапопорта (1965, 1972а,б, 1973а,б, 1980, 1984, 1987, 1989, 1991, 1993, 1996).
Химия или генетика
Многие современные ученые, занимающиеся эволюционными синергетическими проблемами, продолжают упорно думать, что предбиологическая фаза — это химия, из нее будто бы возникли сложные, совершенные и, несомненно, самые антиэнтропийные формы жизни. Ранее П.Гленсдорф и И.Пригожин, в частности, утверждали, что живые организмы в определенной степени можно рассматривать как открытые химические и физические системы. Правда, позже И.Пригожин и И.Стенгерс (2000а) уже сетовали, что трудно понять переход от «химической» истории, когда отдельные молекулы синтезируются и распадаются, к «биологической» истории, когда многообразие путей химических реакций становится решающим образом взаимосвязанным в процессе сохранения новой индивидуальной сущности, и откуда возникает биологическая информация.
Действительно, трудно понять, если не знать, что между химическим субстратом и биологическим миром лежит самая мощная система созидания — дискретный аппарат наследственности, новая система статистики генетической природы, она, хотя и открытая, но отличается от других открытых систем термодинамики тем, что наделена стационарностью, нулевым уровнем. Именно генетика, оторвавшись от химии и заняв уникальное положение в системе природных атомизмов, создала вокруг себя привилегированную протоплазматическую среду с хорошо организованными физико-химическими формами, базирующимися на сниженном уровне энтропии по сравнению с чисто химическими полимерами, которые очень легко переходят от состояний упорядоченных к состояниям неупорядоченным. Именно генетика дала возможность существовать биологическим формам (Рапопорт, 1965, 1984, 1996). Она сделала переход от неживой природы к живой не резким.
Правда, химия передала генетике дискретные микрофизические формы — кварки и нуклоны, валентные электроны и атомы, а также эстафету каталитических форм. Тем не менее химия не достигает высот собственного молекулярного атомизма, потому что все процессы и явления в ней остаются верными II началу термодинамики (Рапопорт, 1984).
Исключение составляют самоорганизующиеся элементарные открытые каталитические системы (ЭОКС). Концепция ЭОКС, лежащая в основе эволюционного катализа, показывает, что особые неравновесные объекты в химии способны к прогрессивной эволюции, приводящей к возникновению жизни (Руденко, 1980, 1995, 2000, 2003). В несколько ином, более развернутом виде эту мысль выразил И.А.Рапопорт (1974): «Внутри химической формы есть ведущие параметры, не позволяющие навсегда замкнуть в химическом состоянии малые новые пики материальных проявлений. По этой причине, хотя нет примеров самопроизвольного развертывания из химии автономного нового комплекса дискретных измерений, такая вероятность все-таки есть. Она требует, однако, очень многого и включает реализацию собственно атомного набора измерений». В другом месте И.А.Рапопорт пишет (1973), что именно «…химический катализ увеличивает вероятность возникновения новой нехимической стихии с новым и богатым элементарным атомизмом». Между тем нельзя исключить, что в системе химического катализа уже могут быть рассеяны предвестники нового атомизма, или он уже существует нацело. И тогда законно ожидать в ближайшем будущем возникновение зачатков (ростков) новых высокоорганизованных живых форм, которые пока еще недоступны нашему воображению.
Генетическая термодинамика
Итак, в генетике, в отличие от химии, господствует дискретность. По мнению И.А.Рапопорта (1978), генетическая субстанция относится к самым совершенным образцам природного атомизма. Она сохраняет высокую упорядоченность, потому что изменения, возникающие в ней, очень часто носят мелкий, незначительный характер.
Если химические полимерные соединения отличаются высокими показателями энтропии, то генетические тела, как и стационарные электроны в атомах, порождают малую энтропию, причем выстраивается следующий ряд: энтропия нуклеотидов < энтропии триплетов < энтропии гена < энтропии хромосомы < энтропии генома. Сравнительно более высокие показатели энтропии у двух последних генетических атомизмов, вероятно, вызвано тем, что они непосредственно связаны с внутриклеточной сферой, где развернут обмен веществ (Рапопорт, 1974).
Работы И.А.Рапопорта (1965, 1972, 1973, 1993) раскрывают суть нового, четвертого закона термодинамики, согласно которому специфический нулевой уровень генного состояния достигается при Т=300 К (±25?С): limS=0, что невозможно для других форм материи. Так, по выражению
T->300K
Нернста-Планка, энтропия однородного твердого тела или жидкого
химического вещества обращается в нуль при абсолютном нуле температуры: lim S=0.
T—>0.
Именно при этой температуре молекулярная упорядоченность перестает влиять на физические явления, открытые энергетические проявления невозможны. При этих условиях молекулы не вступают ни в какие реакции, однако при все более высоких температурах все без исключения виды молекул несут энтропию, иначе говоря, для химических молекул известная положительная энтропия является при 300К обязательной фундаментальной характеристикой — пишет Рапопорт. Что касается генетического субстрата, далее замечает автор, то он располагает дезэнтропической характеристикой (Т>0, -dS) во всем температурном диапозоне, при котором возможна жизнь. Однако выше 52?С меняются генные конфигурации, а за ним следует деградация от dS=0 к dS>0 (этот факт Рапопорт объясняет тем, что на высоком температурном пределе сказывается громоздкость генного состояния, которую уже не могут полностью контролировать ни валентные связи, ни генетическая энергия, ни стационарность), и, наоборот, при понижении температуры в направлении к абсолютному нулю генная форма приближается к уровню Нернста (третий закон термодинамики, которому, как известно, отвечает состояние химических тел, при котором dS=0 зависит от Е=0 и Т=0 К), не переходя в него необратимо. С последующим постепенным повышением температуры от абсолютного нуля до физиологического значения генное состояние полностью восстанавливается.
Мутации — генетический образ хаоса
Мутации — это любые количественные или качественные перемены в наследственных структурах — генах и хромосомах, аналоги термодинамических флуктуаций, источник генетического беспорядка, один из механизмов нарастания генетического и биологического многообразия в природе. Появляясь внезапно, мутации, как и революции, разрушают и созидают, но не уничтожают законы природы. Они сами подчинены им.
Впервые основные положения о мутациях как фундаментальном биологическом явлении были изложены в обширном труде известного голландского ботаника, одного из переоткрывателей менделевских закономерностей Гуго де Фриза «Мутационная теория», вышедшем в 1901 г.:
- Мутации возникают внезапно, без всяких непрерывных переходов, как дискретные изменения признаков
- Новые формы константны, т.е. устойчивы
- Мутации являются качественными изменениями
- Мутации происходят в разных направлениях, они могут быть как полезными, так и вредными.
- Вероятность обнаружения мутаций зависит от числа исследованных особей, проанализированных для обнаружения мутаций
- Одни и те же мутации могут возникать повторно
Красной нитью через работу Гуго де Фриза проходила мысль: «Изменения во внешнем виде организмов возникают в виде скачков. Эти скачки малы, и они наследственны, не образуют никаких переходов, промежуточных ступеней между собой».
Гуго де Фриз считал, что мутации сразу дают начало новым видам, приспособленным к внешней среде, без творческого участия естественного отбора. Этот взгляд противоречил идее Дарвина о том, что внезапные крупные изменения наследственного характера не играют значительной роли в происхождении вида и что видовые признаки возникают путем медленного накопления мелких индивидуальных уклонений. В дальнейшем известные английские ученые Ф.Гальтон и В.Бэтсон выработали примирительную точку зрения: прерывистые изменения, появляющиеся в виде отчетливых новых ступеней являются источником новых наследственных различий, на котором осуществляется естественный отбор.
Предтечей «Мутационной теории» Гуго де Фриза был так называемый неокатастрофизм-мутационизм, включавший в себя концепции немецкого гистолога Келликера и петербургского ботаника С.И.Коржинского о гетерогенезисе, а также идея геолога Зюсса о «перечеканке живых существ», т.е. гигантских превращениях форм на протяжении короткого периода.
Согласно теории гетерогенезиса, изложенной С.И. Коржинским в академической статье (1899 г.), внезапное появление особей, резко отличающихся от родительских форм, происходит вне зависимости от внешних условий жизни. «Когда в силу каких-то неизвестных причин, таинственных процессов, из оплодотворенной яйцеклетки может развиться, вопреки законам наследственности, организм, настолько несходный с родительским, что его как по сумме внешних признаков, так и по способности передавать их по наследству, можно принять за новый вид». С.И.Коржинский считал, что образование новой гетерогенной расы связано с энергией изменчивости, которая, накапливаясь веками, внезапно освобождается и что для объяснения происхождения высших форм из низших необходимо допустить существование в организмах так называемой virtus progressiva (тенденции прогресса, или, говоря на языке синергетики, способности к самоорганизации). Как видно, уже тогда спонтанная наследственная изменчивость не связывалась с воздействиями внешних факторов.
Первым стихийным мутационалистом считается китайский император Кан-Хи (1662-1723), обнаруживший мутант риса, отличавшийся скороспелостью и высоким качеством зерна, названный впоследствии «рисом императора». Самое же название «мутация» впервые было введено садовником Дюшеном в XYIII веке, после установления им
факта внезапного появления земляники с резко измененной формой листьев.
Современная генетика знает, что мутации бывают:
-
Полезные (положительные) и вредные (отрицательные), причем «…во всех случаях, и в природе, и при экспериментальном мутагенезе, отрицательные перемены в наследственных структурах преобладают, но весьма скромно, не более чем на один порядок, а часто значительно меньше».
-
Нейтральные. Они входят в резервный фонд мутаций. Их роль положительна. Предполагается, что они способствуют выживанию организмов в условиях катастрофических изменений условий жизни.
-
Доминантные и рецессивные. Последние возникают в 10-100 раз чаще, чем первые. И, как пишет И.А.Рапопорт, причина этому очевидна: всегда более вероятны мутации, в структуре которых наступают упрощения или даже частичная деградация.
-
Встречаются также мутации, по отношению к которым доминантный «дикий» тип становится рецессивным.
-
Летальные и полулетальные. Первые поражают особей на эмбриональных и постэмбриональных стадиях развития, вторые — лишены морфологических проявлений, но снижают жизнеспособность
-
Соматические и генеративные. Последние возникают в половых клетках и зиготах, они наследуемы
-
Генные, точечные (добавления, дефициты, перестановки на уровне единичных нуклеотидов), хромосомные (перестройки, крупные потери), геномные (кратные увеличения хромосомного множества).
Изучая явление мутаций, современная генетика не ограничивается анализом самопроизвольных (спонтанных) мутационных изменений, общий вес которых, как считает И.А.Рапопорт, достаточен для такого баланса с естественным отбором, чтобы сделать цепь существования живых форм непрерывной и обеспечить их дифференцировку, но очень часто обращается за помощью к орудиям микрофизики и химии органического синтеза.
Спектры индуцированных мутаций выступают как своеобразные экспериментальные ключи, с помощью которых идентифицируются и реформируются гены в хромосомах, подавляется генетический хаос, вызываются сложные изменения в структуре хромосомных материалов, за которыми следуют изменения системы регуляторных генов, восстанавливается топологическая цельность и нормализуются многие внутриклеточные процессы, создаются новые биологические формы, более упорядоченные, устойчивые и продуктивные, и, с точностью наоборот, снижаются показатели не жизненно важных количественных признаков, например, содержания белка и жира в молоке, шерсти у овец, шелка в коконах, которые были чрезмерно развиты и упорядочены в ходе искусственного отбора. Индуцированные (искусственные) мутации ускоряют формообразовательные процессы, укорачивают циклы развития, с их помощью можно вернуть утраченные в ходе филогенетического развития гены из палеонтологического прошлого.
Важно отметить, что именно советские русские генетики, блестящие ученые Н.К.Кольцов (впервые указавший на ошибки генного синтеза в генезисе мутаций), Н.И.Вавилов, Ю.А.Филипченко, С.Г.Навашин, А.П.Владимирский, А.С.Серебровский, Г.Д.Карпеченко еще в первой четверти ХХ века стимулировали изучение проблем индуцированного (искусственного) мутагенеза. Сегодня значение результатов, полученных в области мутационной генетики, для теоретической биологии, эволоюционного учения, селекции и медицины трудно переоценить.
С синергетической точки зрения термодинамическая роль мутаций, как энтропийной силы, направлена на достижение изменений в структуре генетического материала. При этом она не сводится только к деградации генетических структур до химического, молекулярного уровня, разрушению живых систем и их последующей гибели. Напротив, усиление мутабильности — склонности к мутациям — порождает в наследственном аппарате мощный дезэнтропический потенциал, о чем свидетельствует высокая устойчивость положительных и отрицательных перемен, не уступающая начальным, домутационным состояниям (Рапопорт, 1965, 1972). Почти всегда под влиянием мутаций генетическая основа остается неизменной.
Случайность и отбор способствуют постепенному накоплению мутационных, хаотических изменений в наследственных программах. Запас этих изменений обеспечивает живым существам на фенотипическом уровне большую пластичность, гибкость и высокую степень приспособляемости (Берг, 1966). Биологическое развитие невозможно без мутаций. Если живые системы свободны от мутаций, то даже высокая стабильность генома и наличие большого числа упорядоченных, нормально работающих генов, не спасут их от разрушений в периоды резких катастрофических изменений условий жизни.
Действительно, как явствует из нелинейной динамики, системы с хаосом проявляют удивительную пластичность и чутко реагируют на любые внешние воздействия. «Инновационные» возмущения окружающей среды не способны их разрушить. Более того, они сохраняют присущий им тип движения, развития (Дмитриев, 1998). Так, например, поведение сердечной мышцы по своей природе хаотично, и это позволяет ей гибко реагировать на стрессы, изменения физических и эмоциональных нагрузок. Нарушения хаотичности, нелинейности флуктуаций в биении сердца, появление в нем упорядоченности служат важным признаком приближающейся болезни (Дмитриев, 1998, Лоскутов, 2000). Добавим (см. Пригожин, Стенгерс, 2000а), что энцефалограммы больных эпилепсией не демонстрируют хаотической картины, они регулярны.
В одной из рецензий, посвященных философо-психологическому анализу фильма «Матрица», я нашел одно весьма интересное место, которое просто «дышит» нелинейной теорией: » Искусственный Интеллект пытался создать счастливое общество. Первая матрица строилась как идеальный мир, где все люди счастливы. Но люди не приняли программу, и всех пришлось уничтожить. Почему? Человечество не приемлет реальности без мучений и нищеты, для полного счастья людям нужны несчастья! Не может человек ощутить себя счастливым без эталона счастья. Все познается в сравнении». Тут все понятно: ведь линейность обычно измеряется через параметры нелинейности — максимальных отклонений от действительной характеристики. Генетическая стабильность, например, оценивается с помощью частот мутаций. И далее. «Линия жизни человека — это синусоида: взлеты и падения. По синусоиде бьется наше сердце, ритмы мозга тоже синусоидальные. Прямая линия — это смерть. Не потому ли все теории идеального общества исторически терпели неудачу».
Итак, системы, не изменяющие законам своего развития, в которых все абсолютно предопределено, неактивно и нет место случаю, не имеют право на жизнь и существование. Они — ничто, и должны пасть.
Возьмем другой пример. Из истории.
В середине XIX века во Франции на Констанцском соборе, когда поборники строгости нравов жаловались на порочную жизнь пап и вопили о необходимости реформы нравов, один из кардиналов (Пьер де Айи) прогремел им в ответ:» Только сам черт может еще спасти католическую церковь, а вы требуете ангелов. Только воровство еще может спасти собственность, клятвопреступление — религию, незаконнорожденность — семью, а беспорядок — порядок. Синергетическая логика кардинала очевидна. Ясно, речь идет о том, что нарушены законы развития общества и чтобы удержать общество от полного распада, изменить и улучшить его внутреннюю структуру, необходимо нелинейное, катастрофическое решение проблемы. И такое решение есть. Это — привнесение в систему, вставшую на путь дезорганизации и регрессивного развития, порядкообразующих, хаотических элементов (сигналов), способных, вступая в игру, аннигилировать нежелательные процессы и беспорядок, запустить механизм самоорганизации — переход системы в качественно иное состояние, более совершенное, упорядоченное и устойчивое.
Однако вернемся к теме.
Точечные генные мутации, сложные и несложные хромосомные перестройки, мобильные генетические элементы (МГЭ), амплификации (умножение копий ДНК отдельных генов), такие «примеси», как вирусы и плазмиды (элементарные генетические частицы), нуклеотиданалоги, а также внешние влияния (химические и физические факторы), несущие беспорядочные сигналы, непрерывно перетасовывают генетический материал, поддерживают в нем хаотическое движение, нестабильности, увеличивают поток информации и генетическую энтропию. Все это естественным образом создает условия для развертывания процесса самоорганизации.
При этом в геноме даже отрицательные сдвиги не лишены собственного реального положительного вклада, поскольку представляют собой широко воспроизводимые новые, иногда принципиально новые конфигурации (Рапопорт, 1965).
Экспериментальные исследования показали, что комбинирование (синергетический эффект) двух мутантных геномов — а «не лучшего с лучшим» — может приводить к взаимному исправлению недостатков; нежелательные мутации превращаются в положительные приобретения (Шмальгаузен, 1968, Рапопорт, 1996). Только благодаря скрещиванию двух родительских форм, хилых, не блещущих высокой жизнеспособностью, удалось вскрыть в генотипе не только отрицательные стороны, но и формирование крупных положительных генетических ресурсов, повышающих статус генома, о чем свидетельствует жизненный подъем у их потомков (Рапопорт, 1977). С помощью введения генов, обладающих полулетальными и летальными действиями, можно подтолкнуть депрессированные организмы на одну из благоприятных для них траекторий развития, активизировать изменения в пределах генных единиц, что приводит к возможности усиления признаков.
Недавно известный ленинградский ученый, специалист в области клеточной биологии Ю.Б.Вахтин на базе теоретических исследований пришел к выводу, что в генетическом мире, хотя и действует принцип, согласно которому вновь возникшая мутация нарушает нормальное функционирование наследственного аппарата, но наряду с этим заметно возрастает вероятность того, что этот неблагоприятный эффект мутаций будет перекрываться их полезным действием, в частности, их способностью подавлять деятельность внутриклеточных генетических паразитов, названных им, хаоногенами. В «норме» хаоногены представляют собой незаконно размноженные копии молекул ДНК различной величины и сложности, так называемые подвижные генетические элементы. Значительная часть паразитных молекул ДНК выходит в протоплазматическую среду. В ней они претерпевают собственную эгоистическую эволюцию, захватывают и осваивают все пространство, размножаются и изменяются, а по механизму обратных связей отрицательно воздействуют на работу генома.
По образному выражению Ю.Б.Вахтина, хаоногены превращают внутриклеточный Космос в Хаос. Они определяют старение и смерть живых систем, причем не только клеточных популяций и индивидуумов, но и целых популяций организмов и даже видов. Случайные (или индуцированные) мутационные изменения становятся не только единственным способом усовершенствовать геном, но и сделать его недоступным для атак паразитных молекул ДНК. Очищение внутренней среды от семейства генетических паразитов с помощью мутаций должно приводить к замедлению процессов старения (Вахтин, 1998, 2002). Да, живым организмам нередко приходится жертвовать относительной стабильностью генетической информации, чтобы противостоять внутренним и внешним силам разрушения и беспорядка.
Кстати, ранее И.А.Рапопорт (1991) уже указывал на возможность изгнания из хромосом с помощью химических супермутагенов транспозонов (мелких блоков генетического материала), свободное, беспорядочное перемещение которых по генетическому строению вносит хаос в нормальное функционирование генов, усиливает мутационный процесс. В качестве аналогии заметим, что и в твердом кристалле всегда имеется определенная часть не закрепленных ионов, способных хаотически перемещаться.
Проецируя собственные идеи на генетический аспект механизма этногенеза, открытого Л.Н.Гумилевым в истории при анализе процессов становления, расцвета и последующего старения и гибели этносов, Ю.Б.Вахтин заключает, что мутации пассионарности, возникающие, как предположил Л.Н.Гумилев (2000), в результате действия энергии, поступающей на Землю из Космоса, оказывают отрицательное воздействие на фенотип индивидуумов. Пассионарии-мутанты, наделенные избытком свободной энергии, не являются гармоническими натурами, они обладают морфологическими и психическими отклонениями, в глазах окружающих мутанты выглядят странными, нетипичными, «чужими», вызывают к себе негативное отношение у окружающих их гармоничных особей. Сами носители пассионарных генов тоже не испытывают дружеских чувств к слабым, пассивным, бездеятельным, нежелающим расходовать энергию членам угасающего общества. Пассионарии-мутанты — число их невелико — 5-10% — искупают свою «худшесть» активной деятельностью, благодаря которой медленно, но верно закладывается новая, эволюционно прогрессивная этносистема.
Мутации пассионарности очень сильно подавляют внутриклеточную эволюцию генетических паразитов, освобождают геном и клетку в целом от накопившегося мутационного груза. Как считает Ю.Б.Вахтин, в качестве своеобразного яда, уничтожающего паразитов и стерилизующего клетки, выступают генопродукты — новые мутантные белки, синтезируемые пассионарными генами. Этот эффект естественным образом затрагивает не только соматические, но и половые, зародышевые клетки, гарантирующие бессмертие генов и непрерывность жизненного процесса, осуществляющие связь между поколениями.
Итак, многое в современной науке говорит за то, что жизнь без хаоса невозможна. Хаос необходим. Он не просто не противоречит порядку, его добавление в систему устраняет нарушение. Наложение двух хаотических подсистем приводит к возникновению конфликта, после которого система сбрасывает энтропийный груз и становится способной создать новую основу для развития.
В живой природе мутации, обновляя и совершенствуя наследственную информацию, создают благоприятные предпосылки для эволюционного прорыва, программируют новые пути развития биологических форм разных уровней структурной организации — от клетки до биосферы и Разума.
Как когда-то заметил известный немецкий ботаник Э.Страсбургер, для выяснения положения организмов в естественной системе главным критерием является высота развития, достигаемая путем мутаций.
Синергетическое представление генетических систем.
В 1865 году Грегор Мендель — в истории науки первый квантовый биолог — предложил обозначать наследственные задатки (в современной терминологии — гены) буквами латинского алфавита: заглавными — доминантные гены, дающие начало господствующим признакам, способным подавлять в гетерозиготном состоянии развитие альтернативных признаков, а строчными — рецессивные гены, детерминирующие образование уступающих, побежденных признаков. Доминантные и рецессивные гены — различные аллельные (энергетические) состояния одного и того же гена.
В генетическом плане любая живая система — клетка или организм — имеет двойное строение, причем идеальной формой (архетипом) генетического строения является система, состоящая из двух идентичных доминантных генов, расположенных в одном локусе, но в двух разных, гомологичных хромосомах, имеющих, как известно, линейную топологию. Сам факт наличия сочленов одной пары генов в двух разных гомологичных хромосомах уже говорит о том, что наследственная материя имеет дискретную, прерывистую природу.
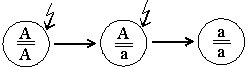
Мутационный процесс чаще всего развивается в направлении от доминантности к рецессивности — А ——> а.
Обычно первая мутация в доминантном гене АА приводит к появлению гетерогенной структуры — Аа, а вторая дает начало новой гомогенной структуре — аа (Рис. 1).
 |
|
Рис.1
|
В генетике и биологии живые системы с такими генетическими структурами называются, соответственно, доминантными гомозиготами, гетерозиготами и рецессивными гомозиготами. Гетерозигота наделена свойствами двух противоположных сторон одной генетической реальности — одновременно доминантной и рецессивной, причем рецессивная мутация (аллель) в гетерозиготном состоянии как бы погружена в виртуальное состояние.
Итак, мутации ведут к утрате чистоты генетического состояния, под их влиянием генная структура эволюционирует от порядка к беспорядку. Согласно точке зрения И.А.Рапопорта, процесс убыли доминантных генов под влиянием спонтанного или индуцированного мутагенеза — явление закономерное и внешне напоминает процесс нарастания энтропии в природе.
Если опереться на аппарат аналогий, то можно увидеть сходство между мутационными превращениями в живых системах и равновесными фазовыми переходами в веществе, связанными с изменениями порядка. То есть ассоциировать доминантную гомозиготу АА, гетерозиготу Аа и рецессивную гомозиготу аа, соответственно, с твердым телом (упорядоченное состояние), жидкостью (менее упорядоченное состояние, система с так называемым промежуточным поведением) и газом (хаотическое состояние). Как известно, энтропия газа > энтропии жидкости > энтропии твердого тела.
Введение еще одной аналогии позволяет сравнить рассматриваемую генетическую триаду с космологическими объектами. Так, доминантную гомозиготу АА можно сравнить со звездами главной последовательности, имеющими нормальную гомогенную структуру, тогда как гетерозиготу Аа — с холодными красными гигантами, имеющими неустойчивую гетерогенную структуру, возникшую в результате скачка плотности на границе ядра и оболочки и, наконец, рецессивную гомозиготу аа можно уподобить белому карлику, энтропированной звезде, внутри которой все ядерные источники энергии полностью истощены.
Недавно известный американский физик, автор теории кварков М.Гелл-Манн сравнил мир кварков и мир блуждающего в ночи ягуара и обнаружил много общего между этими двумя мирами — простым и сложным. Согласно М.Гелл-Манну, кварки как бы призваны символизировать базисные физические законы, которые управляют универсумом и всем веществом в нем…, а ягуар означает сложность окружающего нас мира, в особенности то, как мир проявляет себя в сложных адаптивных системах (цит. по Князевой и Курдюмову, 2000). Однако несколькими годами раньше И.А.Рапопорт (1991) путем проекции генетического строения на физическую кварковую прерывность впервые указал на подобие трех нуклеотидов в триплете и трех кварков в барионе, а затем с учетом параметров прерывности обеих сопоставляемых систем, и на не перестающее нарастание многообразия всех других образцов подобия между микрофизическими и микрогенетическими объектами, связывающими две природные дискретные системы.
Когда скрещиваются между собой доминантные гомозиготы (АА х АА), то всегда во всех последующих поколениях будет воспроизводиться один и тот же «одноцветный» признак, если, конечно, не вмешается мутация. При таком типе скрещивании генетическая энтропия не возрастает. То же справедливо и при скрещивании между собой рецессивных гомозигот (аа х аа). Однако при скрещивании доминантной гомозиготы с рецессивной (АА х аа) в первом поколении возникнет потомство с генетически «неодноцветной», неупорядоченной структурой — Аа. В ней порядок и беспорядок связаны друг с другом. Понятно, при таком варианте скрещивания генетическая энтропия возрастает. Замечательной аналогией здесь выступает заимствованный из статистической физики парадокс Гиббса: при смешивании двух объемов тождественных газов энтропия не увеличивается, тогда как в смешанном объеме, составляемом из двух различных газов, она обязательно возрастает.
Гетерозигота — система на краю хаоса
C позиций термодинамического формализма и синергетики гетерозиготу можно рассматривать как систему генетически неустойчивую, бифуркационную. Ведь в ней теперь одному гену соответствуют два разных аллельных (энергетических) состояний, две степени свободы, а не одна. Это во-первых. Во-вторых, следующая мутация в данном гене может дать начало новому состоянию, которое резко изменит структуру биологического признака.
Гетерозигота — система, занимающая промежуточное положение между двумя полюсами, на одном из которых располагается доминантная гомозигота АА, система с высокой степенью генетической упорядоченности и устойчивости, нулевым (или минимальным) уровнем энтропии, на другом — рецессивная гомозигота аа, система хаотическая, достигшая максимальной степени мутационной энтропии.
Несмотря на то что у гетерозиготных организмов один из источников генетической информации (энергии) ослаблен или даже полностью подавлен доминантным аллелем, они, как правило, сохраняют изотропию, т.е. способность воспроизводить тот же самый упорядоченный признак что и доминантные гомозиготы. Так, например, дикие формы плодовых мушек имеют красную окраску глаз, и гетерозиготные особи все будут одинаково красноглазыми. И хотя считается: то, что внешне не изменилось, не имеет энтропии, это только кажущаяся упорядоченность.
Как утверждает крупнейший генетик ХХ века, основоположник хромосомной теории наследственности Томас Г.Морган (1924), гетерозиготы представляют собой типично промежуточные формы, но с пределами вариации, приближающимися к доминантному (дикому) типу. Поэтому в целом ряде случаев отделить промежуточные формы от диких практически невозможно, но можно легко отличить от рецессивов.
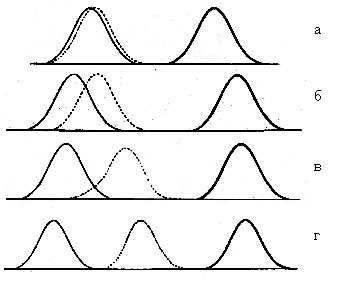
Морган выразил распределение доминантных гомозигот (тонкая), гетерозигот (пунктирная) и рецессивных гомозигот (жирная) в форме вариационных кривых (рис. 2), причем кривые, характеризующие распределения гетерозигот, смещены вправо. Иногда смещения гетерозигот минимальны и при обычных наблюдениях их трудно отличить от доминантной группы (а); в других случаях смещения более значительны (б,в).
Важно также отметить, что вариационные кривые распределения гетерозигот по какому-нибудь признаку могут иметь более «тяжелые» хвосты, уходящие вправо.
Если бы в наших руках было бы что-то наподобие спектрографа, то, вероятно, в фенотипическом спектре такой гетерозиготной системы мы бы увидели гораздо больше частот, оттенков изучаемого признака, чем у доминантных особей.
Морган считает, что, вообще, явлению доминирования признака нельзя придавать слишком большое значение, поскольку многие из гибридных форм оказываются на самом деле истинно промежуточными между родительскими формами. (То есть в гетерозиготе может возникнуть, как говорится, простая наглядная средняя, сочетающая в себе два альтернативных признака — точно, как и в химии, когда сливаются, например, синяя и желтая жидкости можно получить зеленую окраску).
 |
|
Рис. 2
|
Одним из таких примеров служит садовая ночная красавица. Есть две разновидности этого растения — с белыми и красными цветками. При скрещивании между собой они дают растения с промежуточными розовыми цветками (г). Понятно, в данном случае ни одна из окрасок не может быть названа доминантной.
Что стоит за всеми этими смещениями? Качественно новый генетический порядок, возникающий в гетерозиготной системе под влиянием хаотической структуры — рецессивного мутантного аллеля. Который, вовлекаясь в генетические процессы, ограничивает действие оппозитного доминантного аллеля, увеличивает его энтропию. В целом же в гетерозиготе генная система как бы переходит на две скорости, и смешение информации доминантного и рецессивного генов добавляет хаос в структуру биологического признака.