
Как известно (Маркс-Тарлоу, цит. по Князевой, 2001), край хаоса представляет собой динамичную переходную зону между двумя экстремумами: предсказуемым порядком и непредсказуемым хаосом. Сюда, к краю хаоса эволюционируют все живые системы, здесь они остаются гибко балансирующими в критическом состоянии. Только здесь между двумя экстремумами и человек достигает психологического баланса, наилучшим образом психологически вооружен, чтобы иметь дело со странными и непредсказуемыми событиями в жизни (Князева, 2001).
Гетерозиготы можно рассматривать как систему на краю хаоса. Они более приспособлены к изменяющимся условиям окружающей среды, чем гомозиготы. В природных популяциях естественный отбор благоволит к организмам с наследственной неоднородностью, наличием пар, различающихся аллелей, насыщенностью мутациями. Нередко мутации, вредные в гомозиготном состоянии, могут оказаться чрезвычайно полезными в гетерозиготном. При перемене условий окружающей среды и изменении направления отбора резерв мутаций в гетерозиготном состоянии позволяет популяции в более короткие сроки жизни приспособиться за счет изменения генетической структуры, обеспечивает пластичность. Гетерозиготность природных организмов сохраняется очень долго, в особенности, если эта гетерогенность приспособительная (Рапопорт, 1984).
Как реализуется генетический беспорядок?
Допустим, что какой-то доминантный ген в норме детерминировал структуру и функцию одного жизненно важного фермента. Тогда гибель доминантности вследствие двух последовательных мутационных событий неминуемо приведет к смерти всего живого организма. Принцип двух мутаций одного гена в одной клетке в свое время был положен в основы генетической теории старения (Szilard, 1959) и двухмутационной гипотезы канцерогенеза (Knudson, 1971). Согласно одному из положений, сформулированных крупным американским математиком и квантовым физиком Лео Сцилардом, в процессе старения соматическая клетка остается функциональной до тех пор, пока из каждой гомологичной пары генов, по крайней мере, один остается компетентным и активным. Однако эта клетка перестает нормально функционировать, когда оба гомологичных гена окажутся совершенно неспособными к активным действиям. Кнудсон также считает, что в нормальной соматической клетке должны произойти две рецессивные мутации, прежде чем эта клетка станет злокачественной.
Словом, упал в рецессивность — готовься к смерти.
C другой стороны, генетика доставляет множество примеров, свидетельствующих о том, что мутационные сдвиги в структуре гена не обязательно являются источником его необратимого повреждения или гибели. Рецессивные, мутантные гены могут быть активными, способными совершать работу, а следовательно, воспроизводить биологические признаки.
В принципе это не противоречит термодинамике, которая, как известно, допускает возможность нарастания энтропии во Вселенной без угасания физических процессов.
Мутационная энтропия тоже не влияет на генетические процессы, она может только усиливать случайности.
Таким образом, когда мы анализируем динамику развития генетических систем, мы приходим туда, куда уже давно пришли статистические физики, физики открытых систем: энтропия — это всего лишь функция состояния системы, онтометрическое выражение степени ее (системы) неупорядоченности. Словом, средство, с помощью которого измеряется количество беспорядка в системе. Поэтому, если есть какая-то система с большим энтропийным грузом, это еще не означает, что данная система вот-вот разрушится. Тем более, если эта система открытая, живая.
Здесь уместно вспомнить слова Эрвина Шредингера: «Состояние, при котором уже ничего не происходит, физик называет термодинамическим равновесием, т.е. состоянием максимальной энтропии.
Теоретически такое состояние очень часто еще не истинное равновесие, еще не действительный максимум энтропии. Окончательное установление равновесия происходит очень медленно. Оно может потребовать несколько часов, лет или даже столетий». Этот постулат отца квантовой механики представляется очень важным, поскольку указывает на то, что всегда есть какое-то время для проявления случая, флуктуации, способные вывести систему на новую траекторию развития, более совершенную, более прогрессивную, столкнуть ее (систему) с пути эволюции к состоянию мертвой, инертной материи. Понятно, что для разных систем и процессов это время — разное. Для эволюционной катастрофы, в основе которой лежит массовое вымирание огромных групп организмов, оно может занять десятки или даже сотни миллионов лет. А такие деградационные, энтропийные процессы, как угасание и гибель клеточных систем, отдельных индивидуумов, этносов или даже целых цивилизаций? Они тоже растянуты во времени.
Генетическая самоорганизация
Самоорганизация — процесс спонтанного возникновения из энергии хаоса новых, более сложных и упорядоченных структур — самая таинственная и критическая проблема современной науки. Поэтому одной из задач синергетики является изучение сущности этого специального антиэнтропийного процесса, идущего в системах самой различной природы — от физических и физико-химических до экономических и социальных.
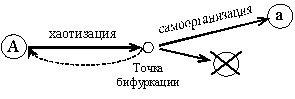
Процессы самоорганизации, конечно, имеют место и в генетическом мире. Чисто теоретически становление и развитие одного из них можно представить себе так. Допустим, что есть какая-то идеальная упорядоченная генная система (доминантный ген А). Предположим далее, что эта система встала на путь самопроизвольной изменчивости (хаотизации) — точечных мутационных изменений. В ходе движения по этому пути система будет отклоняться от закономерной нормы, терять активность и информацию, становиться все более и более неустойчивой. Время идет, энтропия растет. Как только количество мелких квантовых переходов в генной структуре и ее неустойчивость достигнут максимальных критических значений, система войдет в точку бифуркации. И здесь на развилке откроется вероятность для крупного катастрофического скачка: либо возврата системы к исходному начальному состоянию, прежней активности, как говорится, в старые добрые времена, либо вниз — по короткой тупиковой ветви в «энтропический котел молекулярного мира» (разрушительная хаотизация), или же дифференцировки в новом направлении. Движение к новому активному началу, новому энергетическому пику и является, собственно говоря, процессом генетической самоорганизации (созидательная хаотизация) (рис. 3).
Очень возможно, что основу этого антиэнтропийного процесса составляет механизм перегруппировки генетической энергии — перестройки внутренних и межмолекулярных связей в структуре ДНК гена. В результате этих изменений может возникнуть качественно другая генная конфигурация (рецессивный ген а), с новой, ненулевой точкой отсчета роста мутационной энтропии. Правда, в данном случае можно было бы говорить и о нуле отсчета, допустив, что система, которая проходит через слои хаоса и достигает нового устойчивого стационарного состояния, автоматически освобождается от энтропийного груза, забывает свое прошлое. Термодинамика допускает такой произвол.
 |
|
Рис. 3
|
Однако согласно теории Рапопорта, любые мутационные изменения как положительного, так и отрицательного плана должны вноситься в общий энтропический баланс генетического строения. Таково требование мировой энергетической бухгалтерии.
О том, что доминантный ген А перешел по энтропийному мосту в новое квантовое состояние, что на генном уровне (микроуровень) произошла смена структуры (мутация), мы узнаем по количественному или качественному изменениям в структуре биологического признака (макроуровень), когда, например, у фруктовой мушки вместо знакомых красных глаз вдруг видим белые глаза, а у садового душистого горошка вместо привычных гладких горошин обнаруживаем горошины с шероховатой поверхностью. Таким образом, подобно тому, как ключ к атомной структуре лежит в спектре излучаемого атомом света, так и ключ к генной структуре скрыт в фенотипическом спектре. Красные или белые глаза у дрозофилы, гладкие или шероховатые горошины у садового горошка — свидетельство дискретности генного строения. Важно добавить, что частоты мутационных перемен в «атоме» наследственности все же реже квантовых переходов в физическом атоме.
При определенных условиях процесс генетической самоорганизации может и не ограничиться остановкой на «площадке» рецессивности, но по мере нарастания нелинейности развиваться дальше — в направлении к другой, более сложной функциональной упорядоченности в виде нового доминантного гена В (А ——> a ——> B).
Известны также случаи, когда доминантный ген А , минуя состояние рецессивности а, сразу мутирует в ген В (А——>B). Так и звезда после взрыва может «проскочить» состояние белого карлика и сразу превратиться в нейтронную звезду.
Вообще, содержанием мутационного процесса как процесса разупорядочения генного состояния, является не столько реформирование доминантного гена А и возвращение активности и устойчивости в другую дверь, сколько преобразование всей генетической системы: ген ——> иРНК ——> фермент ——> признак. В этой связи я беру на себя смелость предположить, что какие—то силы, действующие не в рамках естественного отбора, исподволь понуждают генетические системы к хаотическим изменениям, способным обеспечить нормальное, устойчивое существование и развитие биологических форм в изменяющихся условиях окружающей среды.
Мутантные, энтропийные гены — очень важные гены
Образование качественно новой полноценной генной структуры — рецессивного гена а — предполагает восстановление генетического веса, способности данной структуры самовоспроизводиться, совершать работу (т.е. быть транскрипционно активной), а следовательно, воспроизводить биологический признак. А если такая мутационная (энтропийная) форма способна создавать биологический признак — не важно, связан ли этот признак с окраской, формой, величиной, структурой или какой-нибудь физиологической особенностью (например, плодовитостью или типами поведения), то выходит, что она не хуже доминантного гена — дикого, чистого, упорядоченного, точно так, как антиводород, состоящий из антипротона и антинейтрона, не хуже водорода, поскольку может производить антиматерию.
Итак, рецессивность — это не упрощение, а усложнение, не слабость, а противоположная доминантность, порядок «наоборот», возникающий как результат неустойчивого бифуркационного процесса.
Из работ И.А.Рапопорта видно, что мутантные рецессивные гены, возникающие в природе спонтанно или под влиянием индуцированного мутагенеза, подчас бывают более пластичные, чем наделенные элитными чертами доминантные гены. Поэтому во многих случаях естественный отбор не отвергает рецессивы, особенно тогда, когда они осуществляют полезный, прогрессивный поворот в строение фермента. Либо участвуют в обрыве энзиматических цепей и колец, осуществляющих метаболические преобразования в клеточных системах, чем, вообщем, содействуют образованию крупных депо полезных продуктов, составляющих критерии продуктивности у культурных растений и животных. Таким образом, не только усложнение, но и упрощение структуры ведет к метаболическому прогрессу.
В определенных условиях система измененных рецессивных генов может проявить себя как специфический, совершенный механизм, удерживающий доминантные гены и детерминируемые ими признаки от избыточной активности и развитости. Более того, роль энтропийных генных единиц со временем может возрастать, когда идеально функционирующие доминантные гены полностью исчерпают внутренние энергетические ресурсы, и станет назревать новый этап изменений в генетической системе. Так и в социальных системах небольшие этнические группы сперва ничем не проявляясь, начинают доминировать в обществе, во всех его сферах.
Другой пример — теперь из биологического мира. На одних островках было обнаружено большое количество насекомых — жуков, бабочек, мух, которые отличались либо полным отсутствием крыльев, либо их крылья были слабо развиты. Согласно старым представлениям, эти островки изначально населялись нормальными крылатыми насекомыми: просто одни летали лучше, другие — хуже, потому что у одних крылья были длиннее, а у других — короче. Сильные бури, которые часто случались на этих островках, уносили отважных летунов в открытое море, и они там погибали. В то время как плохие летуны прижимались к земле, и тем самым сохраняли свою жизнь, преимущества в размножении, оставляли после себя многочисленное потомство. А отбор шел, шел и шел — снова и снова в пользу плохих летунов. Таким образом, в результате длительного действия естественного отбора образовались породы и формы бескрылых насекомых, обладающих высоким коэффициентом приспособления к катастрофическим условиям жизни (Гольдшмидт). Вместе с тем надо подчеркнуть, что информация о длиннокрылости никуда не исчезает, но сохраняется как определенная конфигурация в структуре генетического атомизма. Придет время и генетические детерминанты длиннокрылости вновь выйдут на арену жизни, быть может, в подправленном виде.
Далее в рапопортовской работах находим, что «наборы доминантных генов, которые в основном складывались под влиянием естественного отбора, почти всегда коррелированы с избранными чертами, благоприятными для условий жизни, т.е. существует прямая связь примата доминантности и способности живых существ сопротивляться экстремальным условиям температуры, осадков, ветров, инфекций, вредителей. Между тем и доминантные гены, детерминировавшие на протяжение чудовищных отрезков времени адаптивные признаки, могут создавать тупиковое положение в эволюции генома. И тогда крупномасштабный квантовый скачок к рецессивности открывает вероятность, чтобы осуществить дивергенцию в направлении нового доминантного гена иного строения, контролирующего другую каталитическую функцию. Этот взгляд И.А.Рапопорта на катастрофическую эволюцию генной структуры очень близок тому, о чем пишет А.Хазен, когда анализирует динамику развития социальной системы в условиях катастрофы: система, накопившая огромное количество ошибок, неспособна нормально, устойчиво функционировать, и чтобы выйти из сложившегося тупика, не погибнуть, сохраниться в русле эволюционного (исторического) процесса, необходима революция — хаотизация системы с последующим установлением нового равновесия, как «цели» (аттрактора). И пока ничего лучше этого способа борьбы с принципом максимума производства энтропии Природа не придумала (Хазен, 2000).
Важным резервным материалом для образования новых доминантных генов и, соответственно, новых биокаталитических функций выступают дупликации — линейное следование в хромосоме нескольких одинаковых по структуре и функции доминантных генов (ААААА…). По показателям энтропии избыточные генные единицы мало чем отличаются от активно работающих и важных для жизни генов, и очень часто сохраняются неизменными в своих «гнездах» на протяжении чудовищных отрезков времени (Рапопорт, 1974). И.А.Рапопорт (1996) также показал, что там, где есть довольно большие дупликации, мутабильность понижена в 5 раз по сравнению с соседними недуплицированными районами.
Показано, что этот тип хромосомной мутации не влияет на жизнеспособность клетки и особи в целом. Более того, дуплицированные гены могут усиливать какую-либо функцию организма и способствовать его выживанию. В то же время неразрешенные дупликации часто приводят к онкологическим заболеваниям и другим патологическим проявлениям. Дупликации играют не последнюю роль в эволюции генома и живых систем, селекционных процессах. Из них в результате одного мутационного скачка может возникнуть новый доминантный ген В (А ——> В). Ясно, что такой мутационный переход, называемый дивергентной мутацией, не ведет к потере предшествующего типа доминантной мутации, так как представители ее сохраняются по соседству. С другой стороны, взрыв спонтанной мутабильности в семействе дуплицированных генов в состоянии подавить активность части копий, их способность синтезировать генопродукты — РНК и белки. В генетике такие инактивированные копии называются псевдогенами (Рапопорт, 1974, 1996, Ауэрбах, 1978, Инге-Вечтомов, 1989, Новости науки, 2003).
Другим потенциальным источником образования новых доминантных форм генов, без потерь для других законно существующих генных единиц, является энергия немых генов — генетических тел, находящихся в состоянии термодинамического хаоса. Немые гены не участвуют в создание биологических признаков, поэтому никак — ни косвенно, ни прямо — не взаимодействуют с внешней средой, и закрыты для действия естественного отбора. Немые гены наряду с функционирующими генами и явлением мутаций подчеркивают фундаментальную черту генетического материала — дискретность (Рапопорт, 1996). Мутации вскрывают потенциал немых генов, после чего эти гены становятся способными синтезировать генопродукты и воспроизводить признаки, которые сразу становятся объектом творческой деятельности естественного отбора (Рапопорт,1996). Дивергенция (по Дарвину, увеличение «суммы жизни»), как явление, разводящее признаки у родственных организмов в процессе эволюции и играющее роль в возникновении новых систематических категорий, пользуется потенциалом этой скрытой генетической массы. В геноме эукариотов удельный вес немых, нефункционирующих генов огромен — 90-95% (у растений эта цифра нередко достигает 99%).
Ф.Энгельс в «Диалектике природы» назвал концепцию энтропии — «нелепейшей теорией». Он считал, что излученную теплоту можно будет «вновь использовать». Больцман не был знаком с «Диалектикой природы». Однако согласно его «флуктуационной» гипотезе, проблема обратимости может быть разрешена, а следовательно, мир спасется от тепловой смерти. «Во Вселенной господствует тепловое равновесие и как его следствие — смерть. Но то там, то здесь, в небольших областях — они будут порядка нашего звездного мира (индивидуальные миры) — будут иметь место значительные отступления от термодинамического равновесия».
В хромосомах немые гены и псевдогены, находясь в состоянии теплового хаоса, представляют собой огромную «мертвую» часть генетической энергии, в то время как регуляторные и структурные гены — своеобразную флуктуацию. Мутации, как термодинамические флуктуации генетического происхождения, в состоянии «разговорить» немые гены и псевдогены. И такая вероятность имеется всегда, и она еще более возрастает в условиях экспериментального (искусственного) мутагенеза.
Обратные мутации
С точки зрения временной, Т-симметрии механические и электродинамические процессы всегда могут быть приведены в обратном направлении, т.е., если имеет место процесс А——>В, то с той же вероятностью возможен и обратный процесс В——>А.
Генетический закон не запрещает наследственным структурам возвращение в генетическое прошлое, причем с любой точностью. Правда, вероятность обратных мутаций не очень велика. Обратимые мутационные процессы (а——>А) полностью или частично восстанавливают домутационное состояние, первоначальную последовательность нуклеотидов в структуре ДНК энтропийного гена. Обратные мутации уничтожают последствия прямых мутаций. Способностью вызывать обратные мутации обладают такие генетические мутагены, как транспозоны — одна из форм мобильных генетических элементов (Рапопорт, 1991).
Гомейозисные мутации. Путешествие в прошлое.
И.Пригожин и И.Стенгерс (2000б) утверждают, что время течет в одном направлении, из прошлого в будущее, причем манипулировать временем мы не можем, не можем заставить его идти вспять, в прошлое.
Однако приоткрыть занавес сцены, на которой, как говорится, разыгрывается пьеса прошлого, мы можем. С помощью так называемых гомейозисных мутаций.
Согласно классическим представлениям, гомейозисные (подобные) мутации — это мутации, обращающие эволюционные процессы. Они ведут к восстановлению в существенных чертах каких-то давно нарушенных причинных связей, генов, отомкнувших возникший в другое время запор, кардинальным перестройкам онтогенеза, возврату к структурам далеких по филогенезу предков (Кольцов, 1936, Астауров, 1974, Рапопорт, 1996, Воронцов, 1999).
Так, известно, что однокопытная лошадь произошла от своего трехкопытной лошади-предка. Если кто-то сомневается в этом, — пишет К.А.Тимирязев — то это сомнение снимается появлением уродливостей, например, лошади с дополнительным копытцем. Такая мутантная лошадь была, например, у Александра. Македонского.
В середине 30-х годов академик Б.Л.Астауров открыл и описал у фруктовой мушки Drosophila melanogaster мутацию «tetraptera», в результате которой у этих мушек вместо редуцированной пары крыльев (жужжалец), которые образовались из второй пары крыльев ее эволюционного предка, вновь возникают нормальные крылья. Другими словами, мутация «tetraptera» одним квантовым скачком возвращает представителя двукрылых на миллионы лет назад к древнейшему строению 4-х крылого предка.
Итак, «…природа оставляет орган в виде «следов», чтобы оставаться верной закону неделания скачков» (Ж.Кювье).
Гомейозисные закономерности действуют не только на уровне отдельного, целого органа, но и на другом уровне биологической организации — клеточном.
Наши экспериментальные исследования показали (Захидов и др., 2002, Гопко и др., 2003), что у мутантных ускоренно стареющих мышей образуется сравнительно большое число спермиев с морфологически аномальными головками (ядрами), которые каким-то удивительным образом повторяют конфигурации как нормальных, так и аномальных головок спермиев у крыс, подвергшихся испытанию огнем химического мутагенеза. Этот факт заслуживает особого внимания. Дело в том, что процесс формообразования зрелых мужских половых клеток генетически строго детерминирован и видоспецифичен.
При повышенном мутационном давлении — спонтанном или индуцированном — на наследственные структуры зародышевых, стволовых или развивающихся сперматогенных клеток на терминальных стадиях развития спермиев усиливается морфологический беспорядок, появляется сложная, многообразная картина аномалий ядерных структур.
Образование подобных по геометрическому признаку атипичных спермиев у двух разных, но филогенетически близких видов, говорит о том, что в обоих случаях мы имеем дело с одним и тем же классом гомейозисных, регрессивных мутаций, под влиянием которых современные морфогенные системы уступают место более древней, но общей для мыши и крысы формообразовательной системе. Другими словами мутагенные манипуляции низводят нормальные спермии этих двух видов грызунов к их общему основанию, первобытной форме, состоящей из множества элементарных хаотических структур, представляющих собой то промежуточное звено, которое, вероятно, связывает мышей и крыс с их общим предком, жившим 10 млн. лет тому назад. Итак, между спермиями мышей и крыс существует не только морфологический разрыв, различие, но и глубоко скрытая связь и внутреннее единство.
Наши наблюдения хорошо укладываются в рамки закона гомологических рядов, открытого Н.И.Вавиловым. Согласно этому закону, близкие генетически виды и роды характеризуются сходными рядами наследственной изменчивости. Ряд форм в пределах одного вида дает нам возможность предвидеть нахождение параллельных форм у других видов. У родственных видов, имеющих общее происхождение, возникают и сходные спонтанные и индуцированные мутации по морфологическим, физиологическим и биохимическим признакам. Все это — свидетельство общности происхождения генотипов, и результат мутаций одних и тех же генов. Закон Вавилова не ограничивается только уровнем организмов, но распространяется и на отдельные их структуры (см. Лобашев, 1967, Инге-Вечтомов, 1989).
И последнее в связи с гомейозисными мутациями. Совсем недавно весьма интересное наблюдение о явлении возвратной эволюции сложных структур в естественной природе было сделано американскими исследователями (Michael F.Whiting et al., цит. по С.Мирски, 2003), показавшими, что крылатые виды палочников произошли от бескрылых, которые, в свою очередь, имели предков с крыльями. Авторы убеждены, что возвращение к крыльям — это не результат случайности (действительно, как известно, обращение — процесс «не естественный», он требует воздействия извне). Просто, в течение 50 млн. лет генетические копии ожидали того момента, когда полет стал более благоприятным фактором, чем, например, плодовитость.
«Прошлое еще впереди», — как-то заметила М.Цветаева.
«Одичавший», «развивающийся назад» палочник — знаковая система, первая ласточка переходного катастрофического смешанного времени. Времени, в котором мы уже живем.
Источники использованной литературы
1. Аршинов В.А, Буданов В.Г. Синергетика постижения сложного // Синергетика, Москва-Ижевск, Институт компьютерных исследований, 2003, т.5, с.86-114.
2. Астауров Б.Л. Наследственность и развитие. Избранные труды // М., «Наука», 1974, 359с.
3. Ауэрбах Ш. Проблемы мутагенеза. М., Изд-во «Мир», 463с.
4. Баранцев Р.В. Имманентные проблемы синергетики // Вопросы философии, 2002, №9, с.91-101.
5. Беляев М.И. Милогия и синергетика // Адрес в Интернете http://milogiya.narod.ru/sinergia.htm
6. Берг Р.Л. Играя со смертью // Знание-сила, 1966, №4.
7. Буданов В.Г. Трансдисциплинарное образование, технологии и принципы синергетики // В кн.: Синергетическая парадигма, М., «Традиция-Прогресс», с.284-304.
8. Василькова В.В. Характеристики (атрибуты) порядка и хаоса: от древних космогоний к современной синергетике // Адрес в Интернете:http://lpur.tsu.ru/ Public/art98/a01398.html.
9. Вахтин Ю.Б. Мутации пассионарности Л.Н.Гумилева: возникновение и фенотипическое проявление // 1998, Адрес в Интернете: www.gumilevica.ru.
10. Вахтин Ю.Б. Старение как метагенетический процесс // Цитология, 2002, т.44, с.218-223.
11. Воронцов Н.Н. Развитие эволюционных идей в биологии. М., Прогресс-Традиция, 1999, 640с.
12. Гленсдорф П., Пригожин И. Термодинамическая теория структуры устойчивости и флуктуаций. М., «Мир», 1973, с.280.
13. Гопко А.В., Захидов С.Т., Маршак Т.Л. и др. Генетическая нестабильность мужских. половых клеток у мышей-долгожителей SAMP1, склонных к ускоренному старению// ДАН, 2003, т.392, с.267-270.
14. Гумилев Л.Н. Конец и вновь начало. М., Рольф, 2000, 384 с.
15. Данилов Ю.А. Роль и место синергетики в современной науке // Московский Международный синергетический форум, 1996, Адрес в Интернете: http://www/synergetic/ru/science/index/ pnp/article=dan2.
16. Дмитриев А.С. Детерминированный хаос и информационные технологии // Компьютерра, №47, 1998.
17. Еленин Г.Г. Явления пространственно-временной самоорганизации в системах с многовариантным поведением // Синергетика, Изд-во МГУ, 2000, с.224-242.
18. Захидов С.Т. Принципы подобия между объектами физического и генетического микромира в теоретических трудах И.А.Рапопорта //В кн.: Иосиф Абрамович Рапопорт — ученый, воин, гражданин. Очерки, воспоминания, материалы, Изд-во «Наука», 2001, 335с.
19. Захидов С.Т. Развитие с ускоренным старением // Синергетика, М., Изд-во МГУ, 2003, т.6, с.155-162.
20. Захидов С.Т. Биологическая самоорганизация // там же, с.162-165.
21. Захидов С.Т., Маршак Т.Л., Урываева И.В. и др. Цитогенетические аберрации в клетках печени и сперматогенного эпителия у ускоренно стареющих мышей линий SAMP1 и SAMR1 // Онтогенез, 2002, т.33, с. 444-456.
22. Иванов О.П. Особенности самоорганизации сложных систем в процессе эволюции // Синергетика, Изд-во МГУ, 2000, т.3, с.264-272.
23. Иванов О.П., Малинецкий Г.Г., Рагозин А.Л. Синергетика природных опасностей и рисков // Cинергетика, М., Изд-во МГУ, 2001, т.4, с.57-74.
24. Инге-Вечтомов С.Г. Генетика с основами селекции. М., «Высшая школа», 1989, 591с.
25. Капица С.П., Курдюмов С.П., Малинецкий Г.Г. Синергетика и прогнозы будущего. М., Эдиториал УРСС, 2001, 288с.
26. Климонтович Ю.Д. Шаги к признанию // Знание-сила, 1983, №3.
27. Климонтович Ю.Д. Введение в физику открытых систем. I. // В кн.: Синергетика, М., Изд-во МГУ, 2000а, т.3, с.100-120.
28. Климонтович Ю.Д. Введение в физику открытых систем. II. // там же, с.121-142.
29. Князева Е.Н. Методы нелинейной динамики в когнитивной науке // В кн.: Синергетика, М., Изд-во МГУ, 2001, т.4, с.174-186.
30. Князева Е.Н., Курдюмов С.П. Синергетическое расширение антропного принципа // В кн.: Синергетическая парадигма, М., «Прогресс-Традиция», 2000, с.80-106.
31. Князева Е.Н., Курдюмова С.П. Жизнь неживого с точки зрения синергетики // Синергетика, Изд-во МГУ, 2000, т.3, с.39-60.
32. Курдюмов С.П., Малинецкий Г.Г. Синергетика и системный синтез // В кн.: Новое в синергетике. Взгляд в третье тысячелетие, М., «Наука», 2002, с.3-29.
33. Кольцов Н.К. Организация клетки // Л -М., Изд-во «Биомедгиз», 1936, 652с.
34. Коркоран Э. Упорядоченный хаос // В мире науки, 1991, №10, с.40-41.
35. Лобашев М.Е. Генетика. Л., Изд-во ЛГУ, 1967, 751с.
36. Лоскутов А.Ю. Нелинейная динамика, теория динамического хаоса и синергетика // Компьютерра, 1998, №47.
37. Лоскутов А.Ю. Синергетика и нелинейная динамика: Новые подходы к старым проблемам // Синергетика, 2000, т.3, с.204-224.
38. Малинецкий Г.Г. Синергетика. Король умер. Да здравствует король // Синергетика, Изд-во МГУ, 1998, т.1, с.52-80.
39. Малинецкий Г.Г., Курдюмов С.П. Синергетика и прогноз. Настящее и будущее// В кн.: Новое в синергетике. Взгляд в третье тысячелетие, М., «Наука», 2002 , с.29-58.
40. Мирски С. Возвратная эволюция // В мире науки, 2003, №3, с.10.
41. Морган Томас Гент. Структурные основы наследственности. М-Л, Гос. Изд-во, 1924, 306с.
42. Николис Г., Пригожин И. Познание сложного. М. Эдиториал УРСС. 2003, 344с.
43. Пригожин И., Стенгерс И. Порядок из хаоса. М., Эдиториал УРСС, 2000а, 312с.
44. Пригожин И., Стенгерс И. Время, хаос, квант. М., Эдиториал УРСС, 2000б, 240с.
45. Рапопорт И.А. Микрогенетика. М., «Наука», 1965, 427с.
46. Рапопорт И.А. Модель формирования генетического вещества // В кн.: Химический мутагенез и создание селекционного материала, М., «Наука», 1972, с.13-43.
47. Рапопорт И.А. Сопряжение созидательных процессов на генетическом и селекционном уровнях // там же, с.43-72.
48. Рапопорт И.А. Развитие структуры компакта Дирака в генетическом строении // В кн.: Применение химических мутагенов в сельском хозяйстве и медицине, М., «Наука», 1973, с.7-46.
49. Рапопорт И.А. Природа интеграции и отображения в генетическом атомизме // там же, с.47-76.
50. Рапопорт И.А. Различия в механизме действия основных мутагенов и нуклеотид-аналогов // В кн.: Успехи химического мутагенеза в селекции, М., «Наука», 1974, с.5-28, 29-76.
51. Рапопорт И.А. Определение частоты неизвестных ранее мутаций при опытах по химическому мутагенезу в селекции // В кн.: Химический мутагенез и создание сортов интенсивного типа, М., «Наука», 1977, с.3-36.
52. Рапопорт И.А. Хромосомы в репарационном процессе// В кн.: Химический мутагенез и иммунитет, М., «Наука», 1980, с.3-35.
53. Рапопорт И.А. Действие генетически активных веществ на фенотип и чистота генетического состояния // В кн.:Химический мутагенез в повышении продуктивности сельскохозйственных растений, М., «Наука», 1984, с.3-56.
54. Рапопорт И.А. Значение генетически активных соединений в фенотипической реализации признаков и свойств // В кн.: Химический мутагенез в селекционном процессе, М., «Наука», 1987, с 3-53.
55. Рапопорт И.А. Действие ПАБК в связи с генетической структурой // В кн.: Химические мутагены и пара-аминобензойная кислота в повышении урожайности сельскохозйственных растений, М., «Наука», 1989, с.3-37.
56. Рапопорт И.А. Генетическая дискретность и механизм мутаций// В кн.: Химический мутагенез и проблемы селекции, М., «Наука», 1991, с.3-61.
57. Рапопорт И.А. Две системы прерывности и термодинамическая флуктуация в генетическом строении // Химический мутагенез и задачи сельскохозяйственного производства, М., «Наука», 1993, с.3-24.
58. Рапопорт И.А. Гены, эволюция, селекция. Избранные труды. М., «Наука»,1996, 249с.
59. Режабек Е.Я. Платоновская парадигма и синергетика // Адрес в Интернете:http://public uic.rsu.ru/-smakten/publications/Plato-synergetic.ntm.
60. Руденко А.П. Эволюционная химия и естественно-исторический подход к проблеме происхождения жизни // Журн. ВХО им. Д.И.Менделеева, 1980, т. 25, с.390-404..
61. Руденко А.П. Самоорганизация и прогрессивная эволюция в природных процессах в аспекте концепции эволюционного катализа // Росс. хим. журн., 1995, т.39, с. 55-71.
62. Руденко А.П. Самоорганизация и синергетика // В кн.: Синергетика, М., Изд-во МГУ, 2000, т.3, с.61-99.
63. Руденко А.П. Сложные системы и синергетика // В кн.: Синергетика, Москва-Ижевск, Институт компьютерных исследований, 2003, т.5, с.71-86.
64. Синергетика. М., Изд-во МГУ, 1998, т.1, 256с.; 1999, т.2, 232с.; 2000, т.3, 368с; 2001, т.4, 360с.; 2003а, т.5, 336с.; 2003б, т.6, 200с.
65. Синергетическая парадигма, М., «Прогресс-Традиция», 2000, 536с.
66. Тимирязев К.А. Избранные сочинения. М., Огиз-Сельхозгиз, 1949, т.IY, 474с.
67. Фризъ Гуго де. Мутацiи въ ученiи о наслъдственности // В кн.: Новыя идеи въ бiологiи СПБ, Изд-во «Образованiе», 1914, с.1-54.
68. Хазен А.М. Разум природы и разум человека. М., РИО, 2000, 606с.
69. Хакен Г. Синергетика. М., «Мир», 1980, 400с.
70. Хакен Г. Основные понятия синергетики // В кн.: Синергетическая парадигма, 2000, с.28-55.
71. Чернавский Д.С. Информация, самоорганизация, мышление // Синергетика, Изд-во МГУ, 2000, т.3, с.143-182.
72. Шмальгаузен И.И. Кибернетические вопросы биологии. М., Изд-во «Наука», 1968, 223с.
73. Шредингер Э. Что такое жизнь? С точки зрения физика. М., ИЛ, 1947, 146с.
74. Эйген М. Самоорганизация материи и эволюция биологических макромолекул. М., «Мир», с.216.
75. Knudson A. Mutation and cancer: statistical study of retinoblastoma // PNASA, 1971, v.68, p.820-823.
76. Pearson H. Cancer creates embryo // Nature Science Update, Soc. Neurosci.Ann.Meeting, Florida, 2002
77. Szilard L. On the nature of the aging process // PNASA, 1959, v.45, p.30-45.